Buk 5/95 Eine sexualökonomische Annäherung an den Orgonakkumulator (Teil 1)
Autor: Andreas Frank31 Mai
Bukumatula 5/1995
Eine sexualökonomische Annäherung an den Orgonakkumulator – Teil 1
Günter Hebenstreit:
„Es ist so, als ob der Kasten an der Steckdose hängt…
(…und der andere nicht.” Zitat einer Testperson bei der Orgonakkumulatorstudie auf die Frage, ob sie einen grundsätzlichen Unterschied zwischen der Orgonakkumulatorsitzung und einer Placebokastensitzung bemerkt hätte.)
Mein Interesse für das Leben und Werk Wilhelm Reichs bestand schon zu Beginn meiner Studienzeit. Der Gedanke der Verbindung meiner Neigungen mit den Anforderungen des Diplomstudiums der Psychologie kam bald zum Vorschein. Durch die Arbeit von Gebauer & Müschenich (1987) über die „psychophysiologischen Effekte des Orgonakkumulators“ bekam ich Mut, mich auch an eine derartige experimentelle Überprüfung zu wagen. Die experimentelle Planung und das Setting ist nicht als „orgonomisch“ zu bezeichnen, sondern es ist psychophysiologisch ausgerichtet. Sicherlich mußte ich mich orientieren an dem Umstand, daß diese Arbeit als Diplomarbeit der Studienrichtung Psychologie, also als akademische Abschlußarbeit geplant war. Auch hatte ich zu Beginn der Arbeit noch keine genaue Vorstellung davon, wie sich ein Experiment prinzipiell orgonomisch entwerfen und durchführen lassen könnte.
Andererseits verstand ich diese Arbeit als Überprüfung einer persönlichen Hypothese, inwieweit die wissenschaftlichen Systeme der akademischen Psychologie und der Schulmedizin mit der Reichschen Theorie annäherbar sind. Es war fraglich, ob (und warum) ihre Gegensätze so beschaffen sind, daß sich beide gegenseitig ausschließen, ob sie in Koexistenz miteinander bestehen können, oder ob die Befunde (und dann welche) der Theorien Reichs durchaus mit ihnen in Einklang zu bringen sind. Es besteht die weit verbreitete Meinung unter den „Reichianern“, daß die akademische Psychologie und die Schulmedizin „mechanistisch“ sind. Meist wird daraus ein Werturteil (Etikett) abgeleitet, ohne jedoch die wissenschaftlichen Tatsachen zu beachten bzw. die eigene Meinung auch zu begründen.
Ich hatte die Motivation, diesen Ansatz zu hinterfragen, da ich der Meinung war, daß solch eine hochmütige Art Wissenschaft zu betreiben, nicht dazu benutzt werden sollte, das Hinterfragen der genauen Verhältnisse der „mechanistischen“ und der „reichschen“ Wissenschaft zu verdecken. So war es erforderlich, meine eigene Kompetenz drastisch zu erhöhen und mich gründlichst mit der Sexualökonomie und der Orgontheorie auseinanderzusetzen.
Günter Hebenstreit
Allgemeine biologische Wirkung des Orgonakkumulators
Für die Darstellung der biologischen Wirkung des Orgon-akkumulators (OA) sind einige Grundbemerkungen nötig. Besonders bezüglich der biologischen Wirkungen des Orgonakkumulators existieren Unklarheiten und Brüche in Reichs Theorie. Sie waren auch für die Entwicklung der allgemeinen Fragestellung meiner Diplomarbeit von Bedeutung.
Hielt sich Reich bis zur Zeit seiner skandinavischen Emigration in seinen wissenschaftlichen Arbeiten noch an die gebräuchlichen wissenschaftlichen Termini der einzelwissenschaftlichen Spezialdisziplinen, so änderte sich dies im Laufe seiner amerikanischen Schaffensperiode. Ein wesentlicher Grund dafür waren fehlende oder unzureichende Erklärungsmodelle dieser Spezialdisziplinen. Seit Reich die Strahlungserscheinungen der SAPA-Bione und die Strahlungseffekte des Orgonakkumulators studierte, ging er dazu über, einerseits neue Begriffe für seine Thesen zu verwenden, andererseits gab er üblichen Begriffen einen speziellen sexualökonomischen und orgonomischen Sinngehalt (z.B. Vagotonie, Sympathicotonie, Pulsation, Kontraktion und Expansion, etc.). Sie waren seines Erachtens zutreffender, als jene, welche die Naturwissenschaften zu jener Zeit baten. Ein Beispiel sei hier angeführt:
Reich berichtet nach seiner Ankunft in den USA im Rahmen seiner Bionforschungen über Eigenschaften von bionösen Strukturen, von Einzellern und anderen organismischen Strukturen (Blutkörperchen etc.). Noch in seinem Werk „Die Bione“ orientierte sich Reich eigenen Aussagen zufolge (vgl. Reich 1949a) sehr an einer stofflich-materialistischen Betrachtungsweise Auch zog er die ent entsprechenden Paradigmen, die diese Betrachtungsweise bietet, heran, um Erklärungsansätze für seine Beobachtungen formulieren zu können. In der darauffolgenden Zeit, etwa ab 1940, stehen statt stofflich-materiellen Betrachtungsweisen mehr und mehr bioenergetische Funktionen im Mittelpunkt.
Mit diesem Wandel verschiebt sich die Betonung von den materiellen Komponenten auf Erregungsprozesse. Der Theorie Reichs zufolge bedienen sich letztere nur der stofflich-materiellen Zustände. So suchte Reich (1938) noch stofflich-materielle Gefüge und Veränderungen bei der Untersuchung der Spannungs-Ladungs-Formel (S-L-Formel). Später geht es Reich um die eingehende Untersuchung der Prozesse, die sich durch die S-LFormel ausdrücken, also in erster Linie um Erregungs- und Strömungserscheinungen von lebenden plasmatischen Zellen oder Organismen.
Je mehr er sich mit diesen Erscheinungen auseinandersetzte, desto mehr lösten sich für ihn gewohnte wissenschaftliche Anschauungen und kausale Denkmodelle auf. In den Werken nach den „Bionen“ findet dieser Übergang in Reichs Methodik statt. Reich (1949a) zählt die bioenergetischen Grundeigenschaften aller lebenden Materie am Beispiel der Bionbläschen auf:
„Die Orgonenergiebläschen zeigen die Grundfunktion der lebenden Substanz voll ausgebildet: Attraktion, Erstrahlung, Strahlungsbrücke, Verschmelzung und Durchdringung. Diese Funktionen sind spezifische Eigenschaften der Orgonbläschen, denn Bione, die ihre Orgonladung verloren haben, lassen diese Funktionen vermissen. Diese Funktionen sind also nicht stofflich, sondern energetisch begründet. Sie sind spezifische Orgonfunktionen und haben nichts mit Magnetismus oder Elektrizität zu tun.“ (Reich 1949a, S. 68)
Diese Umstellung hat Konsequenzen: Aus dieser neuen Sichtweise erwachsen neue Möglichkeiten, die z.B. Ola Raknes (1970) für verschiedene Wissenschaftsdisziplinen diskutiert. Sie bringt aber auch für den Forschungsbetrieb einige noch ungelöste Fragen mit sich (z.B. die Frage der Subjektivität im Funktionalismus; vgl. Vittinghoff 1977, Boadella 1980). Graphisch zusammengefaßt lassen sich mehrere Entwicklungsstufen in Reichs Forschungsparadigmen feststellen: Zu der oben mitgeteilten Einführung muß noch ergänzt werden, daß Reich sein Schaffen als Psychoanalytiker begonnen hat und in diesem Zusammenhang erstmals die Orgasmustheorie formulierte:
Psychologie: Betonung des „Quantitativen Faktors“ der Libidotheorie; Gesetze der Spannung und Entspannung in der ersten Formulierung der Orgasmustheorie (vgl. Reich 1927).
Phvsiologie: Gesetze der Spannung und Entspannung in dialektischer Kopplung mit biochemisch-elektrischer Auf- und Entladung von Protoplasma; Modell der „Vegetativen Strömung“ von F. KRAUS (1926, 1927); Vegetatives Nervensystem als ein Funktionsträger des Urgegensatzes von Lust und Angst.
Orgonomie: Gesetze der bioenergetischen Erregung, Erstrahlung, Verschmelzung, Pulsation, Anziehung.
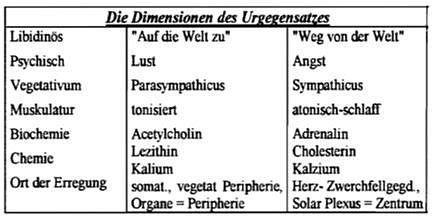
Da sich das Hauptaugenmerk auf der dritten Stufe nicht mehr auf materielle Prozesse selbst konzentriert, wurden die materiellen Veränderungen bzw. Folgeerscheinungen von bioenergetischen Prozessen auch nicht mehr so eingehend von der stofflich-materiellen Seite beschrieben. Im vorliegenden Falle sind dies Begriffe der Vagotonie (s.u.) und der Sympathicotonie, ferner die der Expansion, Kontraktion und Pulsation. Im geplanten Experiment sollen mittels der Erhebung und Messung psychophysiologische Parameter festgestellt werden, ob eine Orgonakkumulatorwirkung tatsächlich reproduzierbar ist.
I. Die vegetativen Wirkungen des Orgonakkumulators
Im Zusammenhang mit klinischen und experimentellen Arbeiten formuliert Reich einen wesentlichen Effekt der Orgonstrahlung: „Die Orgonstrahlung wirkt vagoton …“ (Reich 1949a, S. 182)
Die Strahlung hat dem Zitat zufolge eine Beziehung zum vegetativen Nervensystem. Zu berücksichtigen ist, wie erwähnt, daß Reich die Begriffe sympathisch und parasympathisch (vagisch) in sexualökonomischem Sinne gebraucht. Der normale Gebrauch des Orgonakkumulators läuft darauf hinaus, daß der Patient sich in den mit einer Türe versehenen Orgonakkumulator setzt und darin für ca. 10 bis 60 Minuten verweilt.
„Die Temperaturerhöhung des Organismus ist als eine grundsätzliche Erregungsreaktion der Zellen und des Blutes bekannt. Sie wurde bisher noch nicht verstanden. Diese Temperaturerhöhung weist auf eine Erstrahlung des orgonotischen Körpersystems hin. Genauso wie der Kontakt zweier Bione in eine orgonotische Erstrahlung ausläuft, erstrahlt auch das Blut und das Zellsystem im Kontakt mit dem Orgonfeld des Akkumulators. Dieser Kontakt der beiden Orgonsysteme fiihrt zu einer Steigerung des Orgonenergiewechsels im Organismus, dem mm die belebende Wirkung der Orgontherapie zugeschrieben werden muß. Energiekontakt, Durchdringung Zellerstrahlung und Steigerung des Energiewechsels sind der Reihe nach wesentliche Etappen des Vorgangs.“ (Reich 1949a, S. 322f.)
Demnach besitzt der Orgonakkumulator ein Energiefeld wie ein lebendiger Organismus! Diese Aussage ist insofern bemerkenswert, als sich aus dem bisher Berichteten wohl die Möglichkeit eines solchen energetischen Kontaktes zwischen zwei oder mehreren lebendigen Organismen oder Bionbläschen ergeben kann. Neu ist nun, daß eben auch ein Orgonakkumulator mit seiner speziellen Materialanordnung zu ebensolchem Kontakt geeignet ist (von qualitativen Unterschieden mal abgesehen).
Die anfängliche Annahme, daß der Organismus passiv und unbeteiligt von der Orgonstrahlung durchdrungen wird, war der Röntgen- und Radiumbestrahlung entlehnt. Bei jenen Strahlungsarten handelt es sich um nicht-biologische, körperfremde Energie. Die Orgonstrahlung dagegen stellt eine körpereigene, biologische Energie dar. Sie wird dem Organismus durch die Atmung der Haut, durch Kontakt mit anderen Menschen und Organismen, oder z.B. auch mit der Lunge aus der Atmosphäre „geholt“, oder man bekommt sie von der Sonne zugeführt.
Der Organismus enthält in seinen Zellen und Flüssigkeiten Orgon, er nimmt es auf bzw. strahlt es unausgesetzt ab. Befindet sich nun der Organismus im Orgonakkumulator, so treten zwei orgonotische Systeme in eine funktionelle Beziehung zueinander (vgl. Reich 1949a, S. 318f.). Anhand eines Beispiels aus der Mikrobiologie schildert Reich seine Sichtweise von den energetischen Prozessen zweier in Kontakt tretenden orgonotischen Systeme (das sind im einfachsten Falle Bione mit einer Membranumhüllung oder sonstige ein- und vielzellige Organismen oder rote Blutkörperchen):
„Geraten nun zwei orgonotische Systeme in entsprechende Nähe zueinander, so bilden die beiden Orgonenergiefelder einen energetischen Kontakt. Die nächste Folge dieses orgonotischen Kontakts ist gegenseitige Erregung und Attraktion. Sie äußert sich darin, daß die orgonotischen Systeme einander näherrücken. … Sind die Blutkörperchen nahe genug gerückt, so bildet sich eine Orgonenergiebrücke mit starker Lichtbrechung aus. Jetzt beginnen die beiden biologischen Kerne der orgonotischen Systeme stärker zu strahlen. Wir nennen die Erscheinung die „orgonotische Erstrahlung“.
Sie ist dasselbe, was die Schulbiologie als „mitogenetische Kernstrahlung“ in der Zellteilung beschreibt. [Anmerkung des Autors: mitogenetische Kernstrahlung = Strahlungsphänomen, das bei der Mitose, der Zellteilung, in einer bestimmten Phase auftritt.] Sämtliche fundamentalen bioenergetischen Vorgänge wie Sexualerregung, Orgasmus, Zellverschmelzung und Zellteilung gehen mit hoher bioenergetischen Erregung, also mit orgonotischer Erstrahlung einher. Es handelt sich um starke Energieentbindung in der lebenden Materie.“ (Reich 1949a, S. 319)
Aus der Erstrahlung bzw. „Überlagerung“ der Orgonenergiefelder des Akkumulators und des sich in ihm befindlichen Organismus soll die eigentliche und wesentliche Heilwirkung dieses Gerätes entspringen. Im Falle des oben zitierten Beispiels bildet sich zwischen den beiden orgonotischen Systemen eine Orgonstrahlungsbrücke an den Kontaktflächen. Im Falle des Orgonakkumulators umhüllt das Orgonenergiefeld des unbelebten orgonotischen Systems das Energiefeld des lebenden orgonotischen Systems von allen Seiten (vgl. Reich 1949a, S. 320f.)
„Die wiederholten, durch den Akkumulator hervorgerufenen Erstrahlungen des Organismus äußern sich ja auch darin, daß die roten Blutkörperchen mit der Zeit an biologischer Energie gewinnen und befähigt werden, stärker zu strahlen, praller zu werden, Krebsgewebe zu zersetzen, T-Bazillen zu töten etc., was sie im orgonschwachen Zustand nicht können.“ (Reich 1949a, S. 323)
Soweit die bioenergetischen Funktionen. Eng verbunden mit den Heilwirkungen des Orgonakkumulators (OA) ist das vegetative Nervensystem (vNS). Reich schreibt dem OA eine vagotone Wirkung auf das vNS zu. Die dadurch ausgelöste zentrale Entspannung führt zur peripheren Spannung – eben der „Vagotonie“. Unter diesen Umständen besteht dann wieder die Möglichkeit, die Erregung abzuführen bzw. umzusetzen. Wichtige Erfahrungen über die Wirkungsweise des Orgonakkumulators machte Reich im Rahmen der experimentellen Krebstherapie, bei der die Klienten neben einer Vegetotherapie auch regelmäßig Sitzungen im Orgonakkumulator absolvierten. Zu den biologischen Effekten des Orgonakkumulators meint Reich:
„Das Plasmasystem gibt die chronische Kontraktion auf und beginnt sich zu strecken, vagoton zu erweitern. Mit dieser „plasmatischen Streckung“ geht die Herabsetzung des typischen Krebskrankensclunerzes einher.“ (Reich 1949a, S. 182)
Die einzelnen Reaktionen auf regelmäßigen Gebrauch eines Orgonakkumulators sind individuell verschieden stark ausgeprägt:
„Bei dem einen ist die eine Wirkung, bei dem anderen die andere mehr ausgesprochen. Rötung der Haut, Herabsetzung der Pulsfrequenz, Ausbrechen warmen Schweißes und die subjektiven Empfindungen, daß sich im Körper „etwas lockert“, „anfüllt“ oder „schwillt“ etc. … .“ (Reich 1949a, S. 181)
Reich (1949a) beruft sich auf andere Forscher wie z.B. Burr (dt. Ausgabe 1972), die die Theorie stützen, daß jeder lebende Organismus ein energetisches Feld besitzt, das über dessen materielle Grenzen weit hinausreicht. Bezeichnen jene Forscher dieses Feld als elektromagnetisch, so geht Reich einen Schritt weiter und schreibt die meßbaren Effekte dieses Feldes dem Orgonenergiefeld des Organismus zu.
II. Einige Bemerkungen zur Vagotonie im Sinne Reichs
Reich fand in einer dialektisch materialistischen Ordnung der psychoanalytischen Triebgegensätze, daß aus einem einzigen (Ur-)Gegensatzpaar sämtliche andere Gegensatzpaare abzuleiten sind (vgl. Reich 1949b). Zwar kann hier an dieser Stelle nicht weiter darauf eingegangen werden, jedoch sei auf die „Charakteranalyse“ verwiesen (Kapitel:
„Einige Bemerkungen über den Urkonflikt Bedürfnis – Außenwelt“).
Der Urgegensatz von Sexualität und Angst (bzw. Sexualerregung und Angsterregung) bedient sich der in der Tabelle angeführten somatischen wie psychischen Funktionen. Es soll hervorgehoben werden, daß der „Urgegensatz“ so alt ist, wie das Leben selbst.
Die organbezogenen vegetativen Einzeleffekte, wie sie in schulmedizinischen Büchern zumeist dargestellt werden, müssen hier vernachlässigt werden. Es soll besonders auf die unklare Gebrauchsweise des Wortes „vagoton“ hingewiesen werden. Ein kurzes Beispiel soll zeigen, wie verwirrend manchmal das mit energetischen Bedeutungen erweiterte Begriffsinventar Reichs sein kann. Es geht um die Wirkung der Orgonstrahlung:
Sie [die Orgonenergie; G.H.] lädt lebende Gewebe auf und bedingt Expansion des plasmatischen Systems („Vagotonie“). (Reich 1949a, S. 184)
Dieser Umstand brachte auch unter Reichianem einige Unklarheiten hervor (vgl. Müschenich, 1987). Einerseits geht es um die Aufladung der Gewebe und der Expansion des plasmatischen Systems, zum anderen um den Begriff der Vagotonie. Die Aufladung der Gewebe meint die Steigerung der bioelektrischen Erregbarkeit der Gewebe, die im engen Zusammenhang mit der These der vegetativen Strömung steht. Expansion meint in etwa den sexualökonomischen Grundzustand des Organismus. Er bedeutet, wie sehr der Organismus die Fähigkeit besitzt, in seiner Gesamtheit nach der Spannungs-LadungsFormel zu funktionieren („Es geht mir gut. Ich bin in Kontakt mit mir und meiner Umwelt“).
Kontraktion ist dagegen der Zustand, in dem der Organismus dies kaum oder gar nicht vermag („Ich fühle mich scheußlich, erstarrt, isoliert, etc.“). Reich beschreibt die Symptome der „bioenergetischen“ Kontraktion sehr ausführlich und setzt sie prinzipiell gleich mit einem chronischen Sympathicus Hypertonus. Hier ist eine Verbindung mit der Spannungs-Ladungs-Formel (S-L-Formel) erkennbar. Insofern führt Reichs sexualökonomische Theorie ohne Bruch in die Orgonomie hinein, obwohl Reich nicht sehr ausführlich über die Verbindungen seiner beiden großen Theoriegebäude berichtet.
Daß das physiologische Zustandsbild der (plasmatischen bzw. bioenergetischen) Expansion auf der anderen Seite nicht den Innervationseffekten des Parasympathicus entsprechen, hebt Reich ebenfalls hervor (z.B. Reich 1949a). Damit im vegetativen Bereich die S-L-Formel funktioniert, bedarf es des rhythmischen Wechsels der Sympathicus- und Parasympathicusinnervationen (Reich 1982). Über das Zusammenspiel der Enervationen zu einem einheitlichen System berichtet Reich:
„Die schematische Gegenüberstellung von Vagus und Sympathicus ist in der Tat unrichtig; … Der Wirklichkeit kommt wohl die Vorstellung näher, daß es sich um ein funktionell und morphologisch einheitliches System handelt, das in zwei entgegengesetzten Richtungen funktionieren kann. Die Funktion der Streckung und die der Zusammenziehung wären somit funktionell ein und demselben Organ überlassen.“ (Reich 1982, S. 124f.)
Reich unterscheidet dabei mehrere verschiedene Funktionszustände: Aus einer mittleren Gleichgewichtslage heraus kann sich der Organismus „auf die Welt zu bewegen“, oder sich von ihr „zurückziehen“. In diesem Zustand kann er zwischen den Extrempunkten, der Sympathicotonie bzw. Kontraktion („Angst“) und der Vagotonie bzw. Expansion („Lust“) hin- und herschwingen. Das vegetative Nervensystem ist auch mit der Fähigkeit zur biopsychischen Panzerung ausgestattet. Sie stellt einen „biopathischen Gleichgewichtszustand“ dar, der den Zustand der Expansion als auch den der bewußt erlebten Kontraktion verhindert (biopathisch bedeutet hier, daß der Organismus in seiner Gesamtheit der S-L-Formel zu folgen, ernsthaft beeinträchtigt ist; vgl. Reich 1982, S. 126). So gibt es etwa eine Angstdiarrhöe, die durch Innervation des Vagus zustande kommt; und während des Orgasmus eine lebhafte Motorik. Letztere entspricht einem wiederholten Zusammenziehen und Expandieren und nicht nur bloßer Parasympathicus-Aktivität.
Beziehen sich die tabellarischen Zuordnungen von Vagus zu
Expansion bzw. von Sympathicus zu Kontraktion auf die vegetative
Innervation, so darf nicht außer Acht bleiben, daß Reich jeden Lebensprozeß durch den Wechsel von Expansion und Kontraktion (vermittelt durch Parasympathicus und Sympathicus) bzw. von „auf die Welt zu“ und „in sich zurück“ kennzeichnet. Innerpsychisch macht sich das in Form von Lust- oder Angsterregung bemerkbar. Somit verleiht Reich diesen Begriffen ihre sexualökonomische Bedeutung. Die Sexualökonomie erhält dadurch eine qualitative Komponente, die in der Psychophysiologie vermißt wird.
Zum Verhältnis der drei Stufen zueinander: Reichs Forschungsmethode, der energetische Funktionalismus, ist abgeleitet von der Forschungsmethode (und nicht vom geschichtlichen Konzept) des dialektischen Materialismus. Dieser besagt, daß alles Sein nicht aufgrund von Ideen, Gedanken etc. zustande kommt, sondern aus Gegensätzen entspringt, die in der Materie selbst bzw. in der materiellen Welt angelegt sind (vgl. auch Engels 1952 und 1971). Reich nimmt aufgrund seiner Sichtweise (vgl. in Reich 1934b) an, daß das „psychische“ Sein ebenfalls seinen Ursprung im Materiellen hat und sich aus jenem entwickelt hat.
Die Psyche hat somit einerseits die Funktionen der Physiologie übernommen (s. Tabelle). Andererseits hat sie auch ihre eigenen Gesetzmäßigkeiten entwickelt: Die psychischen Abwehrmechanismen, die es im Somatischen nicht gibt! Derart sind Vegetativ= und Psyche einerseits gleich, andererseits einander entgegengesetzt. Das ist das Modell der funktionellen Identität bei gleichzeitiger Gegensätzlichkeit. Dasselbe gilt nun auch für das Verhältnis der bioenergetischen Funktionen und dem Vegetativum. Auch sie sind funktionell identisch.
Müschenich (1987) berichtet aufgrund der Ergebnisse der Arbeit von Gebauer & Müschenich (1987) über die Unklarheit der Reichschen Definitionen von Vagotonie und Sympathicotonie im Vergleich zu den entsprechenden Definitionen der Schulmedizin:
Zu größeren Mißverständnissen kann allerdings folgende Textstelle führen:
„Aus dem Gesagten geht hervor, daß eine Erhöhung der Körperkerntemperatur in engem Zusammenhang mit vagotonen Vorgängen steht, wenngleich hierbei auch noch andere Vorgänge beteiligt sind“. (Müschenich 1987, S. 68f.)
Nach Brück (1990) kommt die Erhöhung der Hauttemperatur und die erhöhte Durchblutung der Haut durch die Vasodilatation der Gefäße im Bereich der Hautgefäße zustande, die vom überwiegenden Vagotonus hervorgerufen wird. Andererseits wird ein Kerntemperaturanstieg durch eine sympathische Reizung (Grundumsatzsteigerung) und somatische Reaktionen (z.B. Wärmebildung durch Kältezittern der Skelettmuskulatur) hervorgerufen. In diesem Sinne bedeutet eine Kemtemperaturerhöhung einen sympathischen Effekt, und nicht einen vagischen, wie es Reich beschrieb. Müschenich (1987) meint zu diesem Widerspruch:
„Dieses Problem halte ich deswegen für so zentral, weil einerseits Reich Expansion des Lebensapparates stets mit Vagotonie gleichsetzt (und umgekehrt Konstriktion mit Sympathicotonie), andererseits, weil wir in unserer Untersuchung sowohl einen Kerntemperatur- als auch einen Hauttemperaturanstieg während derselben ORAK-Sitzung nachgewiesen haben. Nun können die peripheren Gefäße ja schlecht gleichzeitig kontrahiert und erweitert sein!?“ (Müschenich 1987, S. 69)
Bei dieser Diskussion betrachten Gebauer & Müschenich (1987) die vegetativen Einzelfunktion. Reich betrachtet in seinen Schriften die Begriffe Vagus und Sympathicus unter mehreren Aspekten. Einerseits bedeuten sie für ihn die vegetativen Innervationen mit ihrem gewohnten Sinngehalt (wenn auch mit der Berücksichtigung des Gegensatzes von Peripherie und Zentrum, dem er die Einzelfunktionen zuordnet). Andererseits faßt Reich mit den Begriffen Vagus und Sympathicus zwei antithetische Grundtendenzen des vegetativen Lebens (= Urgegensatz) zusammen, welche schon in den basalen Funktionen der Ionenzusammensetzung des Protoplasmas feststellbar sind.
Die Definitionen weichen voneinander ab und sind miteinander nicht mehr identisch, da einmal das Verhalten von Funktionen von Organen bzw. Organgruppen beschrieben wird. Im zweiten Falle wird der Gesamtorganismus selbst dieser Klassifikation unterzogen. Aus der Erfahrung ist bekannt, daß es so gut wie immer nur ein Überwiegen des einen Teiles des vegetativen Nervensystems gibt. Es gehen die gesamtorganismischen (d.h. sexualökonomischen) Definitionen von „vagoton“ bzw. „sympathicoton“ somit weit über die gewohnten Begriffsdefinitionen hinaus. Das ist ein Dilemma, das sich zum Teil auf Ungenauigkeit zurücicfiihren läßt. Reich hat nur bruchstückhaft die oben erwähnten gesamtorganismischen Definitionen umrissen.
Hinsichtlich der orgonomischen Begriffe bezieht sich die theoretische Grundlage der „Expansion“ bzw. der biologischen Wirkung des Orgonakkumulators nicht auf die Ein zelinnervationen. Vielmehr geht Reich im wesentlichen vom gesamtorganismischen Funktionszustand biophysikalischer Erregung aus. Aufgrund der Arbeit von Gebauer & Müschenich (1987) ist es offensichtlich, daß sich vom aktuellen Zustand der Einzelinnervationen nicht automatisch die gesamtorganismische Situation ableiten läßt: Vagoton im sexualökonomischen Sinne muß nicht unbedingt nur eine vagotone Lage physiologischer Einzelfunktionen bedeuten. Entscheidend ist vielmehr, inwieweit der Organismus (die Person) gesamtorganismisch nach der S-L-Formel funktionieren („pulsieren“) kann, bzw. wie sehr er sich in einem vagoton expandierten Zustand befindet.
Nach Reich funktioniert schon ein einfacher Einzeller nach dem Prinzip der S-L-Formel, also schon lange ehe onto- und phylogenetisch ein organisiertes vegetatives Nervensystem vorhanden ist. Hierin liegen noch große Entwicklungsmöglichkeiten für die Physiologie und Pathophysiologie brach. Eine weitere Diskussion muß auf einen späteren Zeitpunkt aufgeschoben werden.
III. Die Orgonakkumulatorsitzung als gesamtorganismischer Prozeß
Die biologischen Wirkungen des Orgonakkumulators können auch prozeßhaft betrachtet werden: Dem vagotonen Ansatz des Gesamtorganismus geht nach den Beobachtungen Reichs immer eine zentrale Kontraktion (Sympathicus-Aktion) voraus (Reich 1982). Ein Hypertonus des Vagus bedeutet nicht mehr als den ersten Ansatz zur Pulsation entsprechend des S-L-Formel. Im günstigen Falle geht die vagische Expansion in eine pulsierende Schwingung des Organismus über. Hier hat das Phänomen des Bewußtseins große Bedeutung. Nur wenn dieser erste Ansatz zur Pulsation bewußt wahrgenommen wird, scheint sie sich auch in eine Schwingung umzusetzen. Dieser Schwingungsprozeß ist jedoch viel umfassender als die Vagusinnervation selbst.
Heiko Lassek (1982) nimmt an, daß es am Beginn einer Orgonakkumulatorsitzung nach einer initialen Sympathikusaktivität zur eigentlichen therapeutischen Reaktion durch „Stimulierung parasympathischer Zentren“ kommt.
In diesem Sinne ergibt sich folgendes Bild von der Wirkungsweise des Orgonakkumulators: Der Anfangspunkt einer jeden organismischen Reaktion drückt sich in einer „inneren Stauung der Energiebesetzungen“ aus. Das ist immer die erste mögliche Antwort des Individuums auf Reize (Reich 1949b). Der Zustand der zentralen Spannung (vgl. Tabelle 1) drückt sich nun mal in einer sympathischen Innervation aus. Das weitere Schicksal des (gesamtorganismischen) Impulses, der vegetativ als zentrale Stauung auftritt, hängt nun davon ab, ob das Individuum ihn in seinem entsprechenden Ausdruck umsetzen kann (vgl. Reich 194b). Abhängig vom aktuellen Zustand der Panzerung gelingt dies in unterschiedlichem Maße.
Im sexualökonomisch gesunden Zustand entspricht dieser Prozeß einem Expandieren, die (biologische und bioenergetische) Peripherie „lädt sich auf‘. In der Folge verlegt sich die Spannung (Erregung, Energie) in die Peripherie, wo sie sich in Handlung und Verhalten manifestiert. Findet dieser Prozeß nicht statt, so verbleibt die Spannung zentral bestehen, ändert ihre Qualität und macht sich als Unlust bzw. Angst bemerkbar. Bleibt die Erregung an der Peripherie „hängen“ bzw. kann sie nicht adäquat umgesetzt werden, dann ändert sie wiederum ihre Qualität und schlägt in Angsterregung um. Die gestaute Erregung (Angst) wird gewöhnlich zum Opfer der Panzerung und entschwindet weitgehend oder ganz der bewußten Wahrnehmung. Flutet die Erregung wieder ins vegetative Zentrum (Herzbereich, Solar Plexus) zurück, dann kommt es ebenfalls zu einer zentralen Stauung von Erregung.
Lassek (1982) liefert auch einen Beitrag zu dem Diskussionspunkt, ob die Veränderungen durch die Orgonakkumulatorsitzung nun einem erhöhten Sympahicotonus oder einer dominanten Vaguswirkung entspricht:
„Den schwer zugänglichen Daten über die ärztlich therapeutische Anwendung des ORAKs und eigenen Erfahrungen zufolge ist die Wirkung der konzentrierten Orgonenergie auf den menschlichen Organismus zweiphasig zu charakterisieren: Auf eine initiale Phase erhöhter Sympathicusaktivität erfolgt die eigentliche therapeutische Reaktion durch Stimulierung parasympathischer Zentren, die die jedem Akkumulatorbenutzer bekannten Erscheinungen der Wärme, vermehrter Hautdurchblutung, zunehmedde Darmbewegung, Entspannung etc. hervorbringen … “ (Lassek, 1982, S. 60)
Zuerst reagiert der Organismus mit einem erhöhten Sympathicustonus. Erst im Laufe der Sitzung(en) wird dieser abgeschwächt, und der Vagus dominiert schließlich die vegetative Innervation. Ist die Hypothese, daß sich zu Beginn einer ORAKBehandlung eine sympathicotone Reaktion einstellt, welche dann darauffolgend immer mehr zu einer vagischen wird, richtig, dann müßten die Versuchspersonen (Vpn) nach einiger Zeit mehr oder weniger ausgeprägte vagotone Innervationen zeigen.
Auch hier besteht die Schwierigkeit, daß sich die Bedeutungen der Begriffe „vagische bzw. sympathische Innervation“ auf der organisch-vegetativen Ebene von denen auf einer bioenergetischen (orgonomischen) Ebene unterscheiden. Funktionell betrachtet liegt die letztere der ersteren zugrunde. Es gibt schon lange vor der Entwicklung eines vegetativen Nervensystems die energetischen Prozesse, die die gleiche Funktion tragen. Es wäre nötig, die für die Erklärung der eigentlichen therapeutischen Reaktion angeführten vegetativen Nervenzentren in ein orgonomisch-funktionelles Erklärungsmodell einzupassen. Deshalb ist diese These vom logischen Zusammenhang her auf der 2. Stufe im Schema richtig und kann dementsprechend experimentell untersucht werden.
Reich (1949a) gibt den Hinweis, daß im Akkumulator eine plasmatische Veränderung stattfindet. Nach Reichs eigener Sichtweise entsteht vor jeder energetischen Expansion eine energetische Kontraktion:
„Jede Plamaströmung beginnt mit einer zentral anspannenden Kontraktion, die sich in eine vagische Expansion auflöst …“ (Reich 1949a, S. 197)
Dieser Vorgang spiegelt sich auf der Ebene des Vegetativums in einer Erhöhung des Sympathicus-Tonus wider.
Zum Abschluß wird noch einmal hervorgehoben, daß einerseits ein Mangel an genauen Definitionen besteht, andererseits unterliegt die Begriffsbildung der sexualökonomischen Konzepte einer „flexiblen, aber bewußten Handhabung durch den Forscher“. Der Forscher muß sich immer im klaren sein, auf welcher Stufe der Begriffsdefinitionen Reichs er sich befindet. Mit diesem Bewußtsein ist es jedenfalls tragbar, daß eben die sexualökonomischen Begriffe an sich sehr wohl valide sind. Aber ungleich leichter ist dann der Umstand zu tragen, daß aufgrund der Weite des Betrachtungsrahmens bzw. der verschiedenen Stufen (s.o.) sie sich einander widersprechen können, bzw. nicht ganz kongruent sein müssen – und das aus gutem Grund!
Für die vorliegende experimentelle Arbeit zum Orgonakkumulator soll folgendes Konzept angenommen werden: Es wird gemäß Reichs sexualökonomischer Theorie bei den vegetativen Einzelfunktionen zwischen zentralen und peripheren Einzelfunktionen unterschieden (vgl. Reich 1942). Die zentralen Funktionen (Herzgegend, Solar Plexus) scheinen sich so zu verhalten, als wären sie sympathisch innerviert. Gebauer & Müschenich (1987) stellten bereits eine signifikant erhöhte Pulsfrequenz und ebenso eine erhöhte Kemtemperatur während der OA-Sitzung im Vergleich zu einer Kontrollbox fest. Die peripheren Funktionen (Temperatur, Muskelspannung) indes dürften sich entsprechend einem erhöhten Vagotonus verhalten. Das bedeutet unter anderem: Verstärkte Darmperistaltik und intestinale Drüsensekretion, erhöhte periphere Körpertemperatur und des weiteren eine tonisierte Muskulatur mit belebten Bewegungen.
Entsprechend diesem Modell können dann eindeutige Vorhersagen gemacht werden, wie sich die abhängigen physiologischen Variablen während der Orgonakktunulatorsitzung verändern sollten.
Zusammenfassung des Experiments
Im psychophysiologischen Experiment wurden die Auswirkungen des von Wilhelm Reich entwickelten Orgonakkumulators (OA) im Vergleich zu einem optisch identischen Placebokasten (Kontrollkasten; KK; Attrappe) untersucht.
Es war nicht die Absicht dieser Arbeit, die Orgontheorie, d.h. die biophysikalische Theorie einer Lebensenergie zu untersuchen. In erster Linie standen im experimentellen Vorgehen die psychophysiologischen Effekte, die im Orgonakkumulator auftreten sollen, im Mittelpunkt. Dabei wurden die Effekte des Orgonakkumulators unter sexualökonomischen Aspekten betrachtet und diskutiert, d.h. unter dem Blickwinkel der Haushaltung biologischer, vegetativer und psychischer Spannung bzw. Erregung. Die weitere Ausarbeitung der experimentellen Methodik unter diesem Blickwinkel geschah in der Sexualökonomie bisher nur lückenhaft und mußte noch ein Stück weit geleistet werden. Schließlich ist sie die Voraussetzung für eine exakte wissenschaftliche Untersuchung.
Reich formulierte die Grundzüge seiner Orgontheorie in engem Zusammenhang mit Beobachtungen, die er mit einem Gerät mit einer speziellen Materialanordnung anstellte: dem Orgonakkumulator (OA). Dieser OA zeichnet sich durch seinen alternierenden Aufbau von einer oder mehreren Schichten von Metall (Eisenplatte bzw. Stahlwolle) und elektrisch isolierenden Stoffen (hier: Preßspanplatten, Kunst-stoffolie) aus. Zumeist besitzt der Orgonakkumulator eine Würfel- oder Kastenform, aber auch andere Formen sind möglich, wie z.B. ein Orgonrohr oder ein Orgonkissen. Im vorliegenden Versuch besitzt der Orgonakkumulator Kastenform, und ist so groß, daß eine erwachsene Person darin sitzend Platz findet. Der Placebokasten besitzt identisches äußeres und inneres Aussehen bei gleichen Abmessungen.
Die allgemeinen Fragestellungen der vorliegenden Arbeit lassen sich wie folgt formulieren:
Sind die physiologischen und psychologischen Effekte, wie sie in der Literatur im überwiegendem Maße dem Orgonakkumulator zugeschrieben werden, auch im experimentellen Laborversuch nachweisbar? Dabei soll insbesondere die Arbeit von Gebauer & Müschenich (1987) überprüft werden.
Die zweite Fragestellung untersucht den Zusammenhang des psychophysiologischen Konstrukts der autonomen Balance (Eppinger & Hess 1910) mit den Effekten des Orgonakkumulators im Vergleich zur Attrappe.
Die dritte Fragestellung beschäftigt sich mit der Suche nach Personenparametem, die durch eine spezielle Wirkung des Orgonakkumulators zustande kommen_ Lassen sich auf der psychologischen Ebene Merkmale finden, die einen entscheidenden Einfluß auf die Stärke und Art der Reaktionen im Orgonakkumulator haben?
Die vierte Fragestellung setzt sich mit der Kasteninnentemperatur während der Kastensitzungen auseinander.
Im psychophysiologischen Experiment sollten die Auswirkungen des Orgonakkumulators im Vergleich mit einer optisch identischen und wärmetechnisch gleichwertigen Attrappe untersucht werden. Es wurde dabei postuliert, daß der Orgonakkumulator eine vegetativ stimulierende bzw. erregende Wirkung auf die biologische Person hat, die unter anderem über Veränderungen des vegetativen Nervensystems einhergehen. Die zusätzliche Attrappensitzung schien nötig, zumal sonst die Gefahr bestünde, daß jede Art von Veränderung, die sich allgemein in einem solchen engen Kasten einstellt, als „Orgoneffekt“ interpretiert werden kann.
Um der experirnentalpsychologischen Forderung nach Objektivität gerecht zu werden, wurden die Versuchspersonen unter Vorschiebung eines anderen Themas („Auswirkungen elektro-magnetischer Felder auf den menschlichen Organismus“) angeworben. Um eventuellen suggestiven Einflüssen des Versuchsleiters entgegenzuwirken, wurde ein Assistent beauftragt, die Reihenfolge der Kästen randomisiert vorzugeben, so daß der Versuchsleiter über den jeweiligen vorgegebenen Kasten nicht informiert war. Die sogenannte Doppel-Blind-Bedingung konnte in den meisten Fällen eingehalten werden. Dort wo sie nicht eingehalten werden konnte (weil der Assistent nicht verfügbar war), wurde diese Veränderung im experimentellen Versuchsmodus (kurz: EXPMODUS) berücksichtigt.
Weiters wurde das Geschlecht der Versuchspersonen in den Versuchsplan aufgenommen und ihr vegetativer Reaktionstyp (VEGTYP; mit den Idealtypen: Sympathicotoniker und Vagotoniker) nach einem modifizierten Modell von Wenger & Cullen (1972) ermittelt.
Um eine physiologische Baseline zu schaffen, wurden die Versuchspersonen vor den Kastensitzungen für die Dauer von ca. 10 bis 15 Minuten einer Ruhesitzung unterzogen. Danach nahmen sie gleich in einem der beiden Kästen Platz.
Es wurden insgesamt 62 möglichst unkundige Versuchspersonen (Vpn) je einmal in den Orgonakkumulator und einmal in den Placebokasten für ca. 20 bis 30 Minuten gesetzt. Der größere Teil der Stichprobe (44 Versuchspersonen) durchlief im Spätherbst/Winter 1991 in Wien den Versuch (Winter- bzw. Stadtversuch). Eine zweite, kleinere Stichprobe von 18 Vpn wurde im Frühsommer in Niederösterreich untersucht (Sommer- bzw. Landversuch). Es wurden deshalb auch unterschiedliche klimatische Bedingungen in den Versuchsplan aufgenommen (Faktor KLIMA).
Um physiologische Veränderungen zu messen, wurden die folgenden Meßvariablen mittels psychophysiologischer Meßgeräte und modernster Computertechnik erfaßt und aus ihnen die in den Klammem stehenden Parameter rechnerisch abgeleitet: Hautleitfähigkeit (Mittleres SCL-Niveau: SCLX; Maximalwert des SCL pro Sitzung: SCLMAX); die periphere Hauttemperatur (Mittelwert HTX und Maximalwert HTMAX); die Achseltemperatur (mittlere Achseltemperatur ATX und der Maximalwert ATMAX); die Herzrate (Mittelwert und Streuung: PX und PSD; SD steht für standard deviation); die Muskelspannung am Musculus Frontalis (auf der Stirn; Mittelwert EMGFX und Variation EMGFSD); die Kasteninnentemperatur (Mittelwert KTX und Maximalwert KTMAX). Während des Winterversuchs wurde außerdem noch die Muskelspannung auf der Schulter (Mittelwert und Streuung EMGTX und EMGTSD) und im Sommerversuch statt dessen die Fußtemperatur (mit Mittelwert FTX und Maximalwert FTMAX) gemessen.
Außerdem wurde unmittelbar nach jeder Versuchsphase ein Befindlichkeitsfragebogen (Basler Befindlichkeitsfragebogen – BBF) vorgelegt, sowie nach jeder Kastensitzung ein eigens konstruierter Fragebogen zur Erfassung von subjektiven Wahrnehmungen und Veränderungen während der beiden Kastensitzungen.
Weitere unabhängige Variablen waren Skalen verschiedener standardisierter psychologischer Tests: Die 6 Giessen-Test Faktoren, die 3 IPC-Skalen und die 2 STAI-Skalen wurden in einer ca. 20 bis 40 minütigen Pause zwischen den beiden Versuchsdurchgängen (Ruhesitzung + Kastensitzung) erhoben.
Aufgrund methodischer und inhaltlicher Überlegungen wurde beschlossen, für die statistische Auswertung sowohl Absolutwerte, also auch Differenzwerte heranzuziehen: Neben den Absolutwerten jeder einzelnen Versuchsphase wurden auch noch Differenzwerte zwischen der Kastensitzung und der ihr jeweils unmittelbar vorausgegangenen Ruhesitzung berechnet. Letzteres sollte dazu dienen, unabhängiger von unterschiedlichen Aktivierungsniveaus der Versuchspersonen zu sein.
Die Auswertung der Daten erfolgte je nach Erfüllen oder Nicht-Erfüllen der Normalverteilungsvoraussetzung entweder durch eine multifaktorielle Varianzanalyse mit den fünf Kontrollfaktoren und einem 4-fach (Absolutwerte) bzw. 2-fach gestuften Meßwiederholungsfaktor (Differenzwerte Kastensitzung minus Ruhesitzung) oder mittels parameterfreien Tests.
Es folgt nun eine Darstellung wichtiger Ergebnisse bezüglich der untersuchten Variablen. Da die Kontrollvariable REIHENFOLGE oft mit den Meßwerten systematisch kovariiert, wird zunächst das Hauptergebnis bezüglich jedes Meßparameters gemeinsam mit eventuellen signifikanten Einflüssen der REIHENFOLGE dargestellt. Wo es keinen bedeutenden Einfluß der REIHENFOLGE auf den OA gibt (sog. Wechselwirkung – WW), wird das nicht extra erwähnt. Anschließend folgt dann eine kurze Diskussion der Wechselbeziehungen zwischen dem OA mit den anderen vier Kontrollvariablen GESCHLECHT, KLIMA, VEGTYP und EXPMODUS.
Bei der Hautleitfähigkeit zeigte sich, daß das mittlere Hautleitfähigkeitsniveau (SCLX) des OA mit 14,3711S(iemens) signifikant höher liegt als jenes im KK, wo das SCLX der Vpn bei 13,2111S liegt. Ebenso sind die SCL-Maximalwerte (SCLMAX) im OA mit 19,29pS signifikant höher als im Kontrollkasten (KK) mit 17,53g. Betrachtet man die Differenzwerte (relative Veränderungen) des SCL, dann nimmt das SCL von RO LRuhephase vor dem Orgonakkumulator) zum OA nur wenig ab (-0,134).
Dem gegenüber fällt die Verringerung des SCL von RK LRuhephase vor der KontrollIcastensitzung) zum KK mit -2,46 viel stärker ins Gewicht. Die Differenzen unterscheiden sich hoch signifikant voneinander. Auch bei den Differenzwerten der SCLMAX-Werte gibt es prinzipiell das selbe Bild: Nur liegen jetzt die Maximalwerte in den Kästen höher als in den Ruhephasen. Von RO zum OA gibt es eine Zunahme von +3,8 itS, während von RK zu KK der SCLMAX-Wert nur um bescheidene +0,754 zulegte.
In allen vier Fällen (2 Absolutwertvariablen, 2 Differenzwert-variablen) liegen die Werte des OA auf einem höheren vegetativen Spannungs- bzw. Aktivierungslevel. Diese Beobachtungen gehen konform mit der Hypothese, daß während der OA-Sitzung ein zusätzliches Wirkungsmoment feststellbar ist, welches verhindert, daß in der an und für sich reizarmen, grundsätzlich desaktivierenden Kastensitzung die SCL-Werte im Orgonakkumulator genauso tief absinken wie im Kontrollkasten.
Bei der mittleren Muskelspannung auf der Stirn EMGFX wurde festgestellt, daß sie im OA mit 4,0511Volt um ca. 1/3 höher liegt als im KK, wo die Vpn durchschnittlich 2,991.1V erreichen. Das heißt, die zusätzlich durch den OA aufgebaute vegetative Spannung drückt sich in einem signifikant erhöhten Durchschnittsniveau aus. Dieser signifikante Effekt zeigt sich bei den Differenzwerten DEMGFX allerdings nicht: Hier sinkt zwar der Mittelwert des EMG-Frontalis von RO zu OA geringfügig um 0,3211V ab, im Gegensatz dazu sinkt er aber von RK zu KK um 2,4611V, also einem Vielfachen, ab.
Trotz dieses deutlichen Unterschieds ist dieser Wert aber nicht signifikant. Es muß hinzugefügt werden, daß der OA während der Untersuchung das EMG in zweierlei Hinsicht beeinflußt: Einerseits hebt er bei der Vp die absolute Muskelspannung an, andererseits ist der relative Anstieg von RO zu OA größer als von RK zu KK. Bei der Signifikanztestung der Differenzen werden nur die relativen Veränderungen verrechnet, es fällt also ein Teil des OA-Effekts von vornherein aus der Analyse heraus.
Bei den Variations- bzw. Streuungswerten des Stirn-EMG (EMGFSD) hingegen läßt sich weder bei den Absolutwerten, noch bei den Differenzwerten ein für einen der beiden Kästen spezifisches Verhalten erkennen. Die 1,5311V Streuung der EMGFSD des OA stehen einem Wert von 1,4411V im KK gegenüber. Die Differenzwerte (DEMGFSD) bewegen sich unter 0,11-1V und sind aus statistischer und praktischer Sicht gesehen bedeutungslos. Die EMGFSD in ihrer Formulierung als ein Indikator für psychophysiologische Aktiviertheit zeigt also keinen Unterschied zwischen OA und KK. Die Vpn liegen in ihrer psychophysiologischen Aktiviertheit in beiden Kästen nicht unterschiedlich hoch. Das bedeutet, daß die Vpn nicht müder oder wacher, nicht aktivierter oder schlapper aus des Kästen kommen. Sie zeigen in bezug auf OA und KK keinen Unterschied.
Bezüglich der Hauttemperatur fanden sich sehr ähnliche Verläufe zwischen den Parametern HTX und HTMAX, weshalb in der folgenden Zusammenfassung der Ergebnisse zumeist beide Parameter betroffen sind: Aufgrund einer hoch signifikanten Wechselwirkung der beiden Variablen HTX und HTMAX mit der REIHENFOLGE ist fürdie Gesamtstichprobe der Unterschied zwischen OA und KK nicht signifikant. Es unterschieden sich also all die Leute, die zuerst im OA und danach im KK waren von jenen, die zuerst im KK und dann im OA gesessen sind. Erst durch die weitgehende Kontrolle und Ausfilterung dieser Wechselwirkung konnte ein signifikanter Effekt bei der Hauttemperatur nachgewiesen werden.
Dabei betrug für die erste Kastensitzung, wenn sie eine OA-Sitzung war die HTX 33,07°C, wenn sie eine KK-Sitzung war 30,91°C. Für HTMAX liegen die Werte im OA bei 33,94°C, im KK bei 32,07°C, was ebenfalls einer Signifikanz auf dem 5%-Niveau entspricht. Aufgrund der Nachwirkung des OA weit über die Kastensitzung selbst hinaus sind die Phasen Ruhe 2 und Kasten 2 deutlich und signifikant beeinflußt und wurden in dieser Analyse nicht berücksichtigt. Dabei sind die Fingertemperaturen nach der OA-Sitzung im Vergleich zur Zeit nach der ersten KK-Sitzung stark erhöht.
Die Differenzwerte DHTX (D steht für Differenz; sonst die gewohnten Parameterkürzel) und DHTMAX sind unabhängig von der REIHENFOLGE und besitzen auch entsprechende Aussagekraft. Beide Variablen sind bezüglich OA und KK hoch signifikant unterschiedlich. Vom RO zu OA steigt die Durchschnittstemperatur relativ um +2,11°C an, bei den Maximaltemperaturen sogar um +3,23°C. Im Vergleich dazu steigen die Durchschnittswerte von RK zu KK um +0,89°C, die Maximalwerte um +1,85°C an.
Zu den Ergebnissen der Achseltemperaturvariablen ist zu sagen, daß sich die Wahl der Achselhöhle als Meßort für die Kemtemperatur als nicht sehr glücklich herausstellte: Durch nicht vermeidbare Bewegungen der Vpn während der Ruhe- und Kastensitzungen kam es immer wieder vor, daß kühlere Umluft in die Achselhöhle drang. Diese führte dort zu einer mehr oder weniger starken Abkühlung der Achselhöhlentemperatur.
Die Folge davon waren eine hohe Varianz und verzerrte Ergebnisse. Eine weitere Instruktion der Vpn schien allerdings nicht ratsam zu sein, da das Ruhig-Halten bzw. das Anpressen des Armes an die seitliche Brustwand auf Dauer sehr störend und unbequem ist, was unter Umständen auch die anderen Meßvariablen beeinflußt hätte. Die Größenordnung dieses Störeffekts ist wahrscheinlich bis zu 5 Mal so groß wie der in der Literatur
beschriebene OA-Effekt. Aus diesem Grunde wurden keinerlei Unterschiede zwischen OA und KK gefunden.
Bei den Pulsvariablen PX und PSD fanden sich Signifikanzen. Jedoch scheint der Mittelwertsunterschied bei PX zwischen OA und KK von +0,28 Schläge pro Minute im OA ein für die Praxis nicht bedeutsames Ergebnis zu sein. Wie kann das denn nun erklärt werden, daß ein statistisch signifikantes Ergebnis nur wenig Bedeutung hat? Hier sei auf die Wahrscheinlichkeit des ß-Fehlers hingewiesen, der mit steigender Anzahl der durchgeführten statistischen Tests zunehmend größer wird. Der ß-Fehler ist das Komplement zum a-Fehler. Inhaltlich bedeutet er die Wahrscheinlichkeit, daß ein statistischer Test eine Signifikanz anzeigt, obwohl die zu vergleichenden Bedingungen sich tatsächlich zu wenig voneinander unterscheiden -das ist eine Angelegenheit der Wahrscheinlichkeiten, die hier aber nicht weiter diskutiert werden soll.
Die Differenzwerte der beiden Versuchsdurchgänge OA-RO und KK-RK von DPX zeigte immer positive Differenzen (= grundsätzlich mehr Aktivierung in den Kästen). Von RO zu OA nahm die Pulsrate um 1,4 Schläge pro Minute (counts per minute; c.p.m.) zu, von RK zu KK um 1,87 c.p.m. zu, was im Statistiktest nicht signifikant blieb.
Anders liegen die Dinge bei der Pulsfrequenz-Variation PSD: Während im OA die PSD bei 7,13 liegt, ist sie im KK bei 7,61. Dieser Unterschied ist im Mittelwertsvergleich signifikant. Die in der Psychophysiologie als Indikator von Vagotonie verwendete Größe zeigt an, daß im KK eine signifikant größere Vagotonie vorliegt als im OA, obwohl die Wirkungsweise des OA als „vagoton“ propagiert wird. Daß hier eine Ungenauigkeit bzw. eine etwas andere Konzeption der Dimensionen Sympathicotonie und Vagotonie zwischen der Sexualökonomie und der (Psycho-) Physiologie vorliegt, wurde bereits ausführlich dargestellt.
Aufgrund der aus der Versuchssituation entspringenden Umstände (enger Kasten, weitgehende Einschränkung der Bewegung etc.) war zu erwarten, daß der postulierte Effekt des OA (Aufbau zusätzlicher vegetativer Spannung bzw. Erregung) ein im Vergleich zum KK höherer Sympathicustonus sein würde, bzw. daß sich beim PSD der höhere Sympathicotonus derart ausdrücken würde.
Die Differenzwerte OA-RO und KK-RK bleiben nicht signifikant. Sie zeigen zu geringe Unterschiede.
Zu erwähnen bleibt wiederum eine Wechselwirkung der REIHENFOLGE mit den beiden Absolutwertvariablen PX und PSD. Es konnte bei PX gezeigt werden, daß nach der OA-Sitzung in der Phase „Kasten 1“ die Mittelwerte der Pulsrate von 74,9 auf 70,6 c.p.m. gesunken sind, was einem vagoton stimulierten Zustand entspricht. Dieser Effekt bleibt individuell verschieden für einige Zeit beobachtbar (ca. 30 bis 60 Minuten), bevor er wieder verschwindet. Der WW-Effekt von PSD ist auf die Phasen RO und OA beschränkt und scheint sich zu einem guten Teil aus der unterschiedlichen Verlaufsdynamik der beiden REIHENFOLGE-Stichproben zu ergeben: Während sich bei jenen Vpn, die zuerst in den KK und dann in den OA gingen, sich im Laufe der Ruhephase 1 (7,07 c.p.m.), Kastensitzung 1 (7,38) und Ruhephase 2 (7,93) die PSD kontinuierlich erhöhte, und dann bei Phase Kasten 2 (= OA) einen Knick in Richtung Sympathicotonus machte (7,03 c.p.m.), findet sich eine Zick-Zack-Bewegung bei der Stichprobe, die zuerst in den OA und dann in den KK ging. Von der Ruhephase 1 zur Kastensitzung 1 gibt es einen Anstieg der PSD von 6,47 auf 7,22 c.p.m. (= OA), bevor sie wieder in Ruhephase 2 auf 7,0 c.p.m. zurückgeht, um dann wieder in der zweiten Kastensitzung (KK) auf 7,81 c.p.m. anzusteigen.
Obwohl eine weitere Analyse von PX und PSD durchgeführt wurde, die eine Ausschaltung des REIHENFOLGE-Effekts verfolgte, brachte sie in bezug auf die Gesamtstichprobe keine weiteren signifikanten Ergebnisse. Jedenfalls konnten Nachwirkungen der OA-Sitzung über die Zeit der Kastensitzung selbst nachgewiesen werden, was für zukünftige Untersuchungen von großer Wichtigkeit ist.
Die Analyse der Variable der Schultermuskelspannung EMGTX brachte als Ergebnis, daß im OA die Muskelspannung signifikant höher ist als im KK. Liegt die EMGTX im OA bei 3,881.iVolt, so ist sie im KK nur bei 3,25 Bei den Differenzwerten trat eine starke WW mit der REIHENFOLGE auf. Nach der Korrektur der WW trat eine signifikant stärkere Verringerung bzw. relative Verringerung der Muskelspannung von RK zu KK im Vergleich zu OA-RO auf. Im OA sind bei jenen Vpn, die zuerst im KK und dann im OA gesessen sind, prinzipiell keine OA-spezifische Nachwirkungen feststellbar. Deshalb wurden nur diese Vpn zur weiteren Analyse herangezogen.
Von RO zum OA kam es nur zu einem geringfügigen Rückgang der Schultermuskelspannung (3,92-4,17gV= -0,25 ktV). Im Vergleich dazu ist die Differenz KK-RK größer: 2,6311V – 4,92 1.1V = -2,29 1.1.V. Diese Differenzen sind entsprechend den Angaben des statistischen Tests (U-Test) signifikant voneinander unterschieden. Das bedeutet entsprechend der Theorie eine vermeinte vegetative Erregung der Muskelspannung (und in der Sexualökonomie wird die Muskulatur auch als vegetatives Organ gesehen), die sich in einem erhöhten Tonus ausdrückt.
Im Hinblick auf die Muskelsparmungsvariation EMGFSD und DEMGFSD sei hier nur festgestellt, daß diese als Indikatoren für psychophysiologische Aktiviertheit in bezug auf OA und KK keinerlei Unterschiede aufzeigen. Vom Standpunkt des Konzepts der psychophysiologischen (bzw. muskulären) Aktivierung gibt es keinen Unterschied zwischen OA und KK.
Bei der Fußtemperatur und den Absolutwertvariablen FTX und FTMAX fand sich wie schon bei den anderen Körpertemperaturen eine starke Wechselwirkung bezüglich der REIHENFOLGE. Weder vor der Korrektur der Wechselwirkung, noch nach dieser konnten hier statistisch signifikante Effekte zwischen OA und KK festgestellt werden. Bei den Differenzwerten DFTX und DFTMAX, die frei von einer solchen störenden Beeinflussung sind, konnten unterschiedliche relative Temperaturrückgänge beobachtet werden: Während bei FTX von RO zu OA die Temperatur um -0,33°C zurückging, fiel sie von RK zu KK um -0,91°C.
Bei FTMAX blieb die Temperatur von RO zu OA gleich, während sie von RK zu KK um -0,58°C fiel. Der verringerte Temperaturabfall während der OA-Sitzung läßt sich insofern als „vagotoner“ Effekt deuten, als die Vp durch das ruhige Sitzen und der Unmöglichkeit der Bewegung der Füße nur geringfügig an Temperatur verlor, während im KK diese Veränderung ungleich stärker ausfiel. Trotzdem gingen allgemein die Fußtemperaturen – auch im OA – zurück!
Beim Befindlichkeitsfragebogen als vorletzte abhängige Variable wurden ebenfalls Absolutwerte als auch Differenzwerte zwischen den vier Versuchsphasen berechnet. Als Hauptergebnis läßt sich feststellen, daß im Versuchsdurchgang mit dem OA die Befindlichkeit (Summenscore; SUMSCORE) niedriger ist, als in jenem mit dem KK. Im KK ist sie mit 80,29 am allerhöchsten, während das SUMSCORE im OA mit 78,19 am tiefsten liegt. Dieser Unterschied von 2,1 Punkten ist signifikant Er verdient – obwohl nicht sehr groß – aber trotzdem Beachtung, da die Streubreite des SUMSCORE sehr hoch ist. Der Erfinder dieses Testinstruments, Hobi (1985), gibt jedenfalls keine Zahlen bezüglich relevanter Unterschiede im BBF an.
Generell läßt sich ableiten, daß die im OA ablaufenden Prozesse die Vpn soweit beeinflussen, daß sie sich in ihrer Befindlichkeit eher eingeschränkt fühlen. Dies könnte einerseits auf die psychophysiologischen Einwirkungen zurückzuführen sein. Auf der anderen Seite muß auf die mit dem Versuch verbundenen Fehlinstruktionen bzw. das Verschweigen des wahren Themas hingewiesen werden. Denn letzten Endes ist nicht abzuschätzen, inwieweit solche, für die Untersuchung notwendigen Rahmenbedingungen, auch tatsächlich in die gewünschte Richtung (Objektivität, Qualität der Daten) wirken. Nicht zuletzt ist die Organempfindung – wenn sie trainiert ist! – in der Sexualökonomie und der Orgonomie prinzipiell einem orgon-sensiblen Meßgerät gleichzustellen und gleichstellbar. Häufig wird aber gerade diese Seite im psychophysiologischen Experiment vernachlässigt bzw. stiefmütterlich behandelt. Hier scheint sich ein großer Graben zwischen den Thesen der Sexualökonomie und der akademischen Psychologie aufzutun.
Bezüglich der Differenzwerte ist sagen, daß diese nur gering und nicht signifikant sind: Von RO zu OA nimmt das DSUMSCORE um 0,47 Punkte ab, von RK zu KK nimmt es um 0,65 Punkte zu. Wie man sieht: da gab es zu geringe Differenzen!
Zuletzt soll noch auf die WW der SUMSCORE-Daten mit der REIHENFOLGE hingewiesen werden. Sie zeigt, daß die SUMSCORE-Werte im KK dann am höchsten sind, wenn die OA-Sitzung vorangegangen ist (77,9 im OA zu 82,9 im KK). Umgekehrt sind die Werte im OA noch niedriger, wenn die KK-Sitzung vorausgegangen ist. Dann haben die Vpn im KK ein SUMSCORE von 79,4, während es im OA auf 77,3 absinkt. Mit einer gewissen Vorsicht läßt sich der hohe SUMSCORE-Wert im KK als 2. Kastensitzung (82,9 Punkte) auf eine Nachwirkung des OA interpretieren, da eine solche sich weder bei der anderen Versuchsgruppe mit der REIHENFOLGE: 1.KK – 2.0A, noch insgesamt ein zeitlicher Effekt erkennen läßt. Die OA-Sitzung wäre übertrieben gesprochen eine bio-physikalische Belastungssituation, während nachher die Früchte der „harten Arbeit“ geerntet werden. Es steht auch zur Frage, ob nicht die Vpn anders reagiert hätten, wenn sie über den Sachverhalt der beiden Kästen informiert worden wären und sie z.B. auch „raten“ könnten, was nun der „richtige“ Kasten ist …
Als letzte abhängige Variable wurde die Kasteninnentemperatur aufgezeichnet: Hier zeigen sich die absolut deutlichsten Ergebnisse von allen bisherigen Variablen, eventuell die mittlere Stirn-Muskelspannung ausgenommen: Durchschnitts- und Maximalwert sind einander sehr ähnlich im Verlauf, weshalb das nun Folgende für beide Parameter gilt: KTX und KTMAX erreichen im OA 24,9 bzw. 25,43°C, während ihre Werte im KK mit 23,85 und 24,35°C (jeweils KTX und KTMAX) deutlich tiefer liegen. Diese Mittelwerte unterscheiden sich hoch signifikant voneinander. Im OA „heizen“ die Vpn den Kasteninnenraum stärker auf als im KK.
Gerade diese Fragestellung grenzt wie keine andere an physikalische Grundlagen, zumal zwar übereinstimmend in der Literatur behauptet wird, daß der OA aufgrund seiner Eisenblech-Innenwände kühler sein müßte als der KK. Andererseits fehlen bis heute Studien zur Problematik des Vergleichs einer mit Eisenblech ausgekleideten Box mit einer Box, die Holzinnenwände besitzt. Und dies, obwohl im nicht-humanen Bereich gerade auf diesem Sektor die meisten Arbeiten auf dem Gebiet der Orgonomie vorliegen.
Die Differenzweite zeigen folgendes Bild: Von RO (RO ist hier die Raumtemperatur) zu OA steigt die Temperatur relativ um 1,68°C bezüglich KTX, und um 2,17°C bezüglich KTMAX an. Im Vergleich dazu ist der Anstieg von KTX bei RK zu KK mit 0,79°C bescheidener ausgefallen, ebenso bei KTMAX: 1,24°C. Beide Variablen erweisen sich im Vergleich der relativen Temperaturzunahmen zwischen OA und KK als hoch signifikant.
Fortsetzung in BUKUMATULA 6/95.
Die Diplomarbeit wurde als Abschlußarbeitfiir die Studienrichtung Psychologie an der Wiener Universität durchgeführt, von Univ. Prof. Erich Vanecek betreut und mit Sehr gut benotet. Bis auf weiteres ist sie im Buchhandel nicht erhältlich.
Eine überarbeitete Version der Diplomarbeit umfaßt ca. 360 Seiten und ist ab Mitte November um 650 öS beim Autor direkt zu beziehen (bei Postzustellung + 33 öS Versandspesen im Inland; Ausland + 130 öS). Nach Einzahlung der o.g. Beträge per Erlagschein auf das Konto der Raiffeisenbank Wr. Neustadt: BLZ: 32937, Nr. 431.379 (Konto: Günter Hebenstreit) und einer kurzen telefonischen Nachricht erhalten sie die Arbeit gebunden im DIN A4 Format zugesandt.
Postadresse:
Mag. Günter Hebenstreit
An der Liesing 2/21/2
1230 Wien
Tel.: 818 70 53.